

��䵱��ʱ��10��2��11:30�������˹�¸��Ħ������˹��ҽѧԺ��ŵ��������ѧ�Լ�ҽѧίԱ����������˹?�������ThomasPerlmann������������ý���Ļ�����ǽܸ���?������Jeffrey C.Hall��, ���˶�?��˹��ʲ��MichaelRosbash�� �����˶�?�Michael W.Young�����Խ����������й������ӷ��ӻ��Ʒ���ķ��֡�
ŵ��������ѧ��ҽѧ���Ǹ����ѹʵ���仯ѧ��ŵ�����������������ģ�Ŀ�����ڱ���������ѧ��ҽѧ������Խ�����ߡ�
�����������������ӷ����о� ������Ƚ����ŵ��

?2017��ŵ��������ѧ��ҽѧ������ܸ���?������Jeffrey C Hall�������˶�?��˹��ϣ��MichaelRosbash�������˶�?�Michael W Young����
��λ��ѧ�ҵĻ������ǣ�
���ֿ�����ҹ���ɵķ��ӻ��ơ�
���˼��
�ܸ���.����(Jeffrey C. Hall)
�ܸ���.����(Jeffrey C.Hall)�������Ŵ�ѧ�ң�1945�����������ŦԼ��1971��������ͼ��ʢ�ٴ�ѧ��ò�ʿѧλ��1971����1973���ڼ�������ѧԺ���β�ʿ���о�Ա��1974�꣬�������˲�����˹��ѧ�ν�ְ���ܸ���.�����������ݡ�
���˶�?��˹��ʲ��Michael Rosbash��
���˶�?��˹��ʲ��MichaelRosbash���������Ŵ�ѧ�ң�1944���������������˹�ǡ�����1970�����������ŵ���ʡ������ѧ��ʿѧλ�������ո����İ�������ѧ�������격ʿ���о���1974���������������ֶ�ɪķ�IJ�����˹��ѧ��ְ��
���˶�. �Michael W. Young��
���˶�. �Michael W.Young���������Ŵ�ѧ�ң�1949������������������ܡ�1975�꣬���ڰ�˹���ĵ¿���˹��ѧ��University ofTexas������˲�ʿѧλ����1975��1977��䣬����PaloAlto��˹̹����ѧ���β�ʿ���о�Ա��1978�������ŦԼ����˷��մ�ѧ��RockefellerUniversity����ְ��
? ����������������������ɵ���
? ������һ���еIJ�ͬ�����Ҳ�������ӵ���
��֪ʶ���ӡ�����������ʱ4��д�����ֳ�����Ƚ�������ӵķ����о���ʷ����ʮһ�������λ���߷������������á�
�����������������ӵķ����о�
�ģ����㣨��֪ʶ���ӡ����ࡢ������ѧ��ϯ���ڣ�
��
���������ڶ���翣�
������������ʵ�ڡ�
��������Ѱ��������
������������������
���о������ӻ���Ĺ����У���������Ԥ���ܷ�ȡ��ͻ�ƣ��º���������������Գɹ����������ǽ�����������Ȼ�������ر���á�
�����������������ܶ���ʼ�Ľ��ɣ���������֪�ģ����������ҹ����ֲ��Ĵ�����ʵ���������Ľ�24Сʱ��ҹ���ɣ�circadianrhythm���ǵ��͵�������֮һ��
�����������Ƽ���ʵ����Ȼ��������֪�����տ�����̫��������ÿ��̫���ȳ��������տ���ת�������տ��ȳ��������̫�������١�
�����ӵı����������֣��о������ס������õ����������о����������ţ�1971��ӹ�Ӭ��һ��������������������ӵĻ����о���23���ŷ��ֲ��鶯���һ�������ӻ����ͻ�䣬26�����ȷ������������ӻ������Ӭ�����ƣ�30���ŷ���ͬ��Ӭһ���Ļ���Ҳ�������������ӡ�
���������ӵ����ڻ�������һ��һ������ķ��ֺ��о��������ʣ��ӹ�Ӭ���˴���ͬ��һ�����������ӵĻ������DZ���ĵ����ʺ������£������Եص���ϸ���ڵĻ���ת¼���������˸�����ģʽ���������¶ȵ��������Э�����Ӷ���Ӧ�ڵ�����ת�Ľ�24Сʱ���ɡ�
�о�����������Ҫ����λ��ѧ�Ҷ���������������λ�о���Ӭ��һλ�о�С��������ѧԺ��SeymourBenzer��1921-2007����RonaldKonopka��1947-2015�����������ӵĻ����о������ֵ�һ�������ӻ���period��per���������Ŷӿ�¡per������˷��մ�ѧ��MichaelYoung��1949-���쵼��ʵ���ң�Brandeis��ѧ��Michael Rosbash��1944-����JeffreyHall��1945-������ʵ���Һ������Ŷӣ����Ƕ������ֵ��������ӵĸ���������������ӻ����о���ͻ�����Ե�ʱ������������ѧ����������ҽѧ���ĵ�JosephS Takahashi������,1951-����
һ�ٶ���������ѧ�������о���Ӭ����1933��1947��1995��2011���Ĵ�ŵ���������������⣬�����ӵ��о����ù�Ӭ�ٶ�������Ŀ��
�����ӵ��о����̣����ֹ���Ȥ�����εĹ��£�
�����ӵ��о��Ƿ��սᣬû���ش����⡢ֻʣϸ���ˣ�
�������о�����ʮ�꣬Ϊ��������δ������Ч��Ӧ�óɹ���
��Ϊ�о��ķ�ʽ�仯
�����ӿ��Կ���һ����Ϊģʽ��
��Ϊ�ǿ��Ա���������������о������۲쵽�Ķ������ڶ������֡���Ϊ�о��ķ�ʽ��1960����ǰ�Ƚϼ���������Ϊ����
���о�����ϵͳ����1904��ŵ���Ķ�����ѧ�Ұ����Ivan Pavlov,1849-1936��������ȴ������������о���Ϊһ��������֪��������ϲ����ʳ��������Һ����ԭ�����������յ���������Һ������������ʳ����ͬʱ���ֺ����������������յ���������Һ��˵�������̼���ʳ��̼�֮����Խ�����ϵ��������ʳ��Ϊ�������̼�������Ϊ�����̼����������̼��ķ�ӦΪ�������䡣���ּ���Ϊѧʵ����20���ͳ���Ϊѧ����Ҫ��չ��
��Ϊѧ��������ѧ��������ѧ��أ�Ҳ������ѧ�ܽ�����������Ϊ���忪����John B.Watson��1878-1958����1913��������Ϊѧ���������ԡ�������˺Ͷ������Ϊ���ܼ��ɺ��쾭�������졣1920������11���µ��к�Albert��ʵ�飬��ͼ˵��ֻҪ����̼��������Ŀ־���Ϊ�����������µ�������������ż������������ù����������ﶼ�ޣ���Ϊ��������������������Ϊ��
������������ѧ��B. F.Skinner��1904-1990���ƹ���Ϊ���壬������ж������һ��������Ϊ���ɺ���̼������죬�Ǽ�����Ϊ����Ĵ������������˾����ʵ����ƣ��������������䡣��������������ֻ������ķ�Ӧ�������䶯����Skinner�ø�����ʵ�飬�ڸ���װ����ť��������һ��ť��õ��ԵĽ��ͣ�����һť����һ��ʱ����ӻᾭ��ȥ���н�ť�����������������������������Ϊ���ֳ���������������Һ���ֱ�����Ӧ����Ϊ�������������䣬��������Ϊ�о�ѧϰ�������Ҫģ�͡�
1973��ŵ��������ѧ��ҽѧ�������о���Ϊ�Ŀ�ѧ�ң��µ�����Konrad Lorenz ��1903-1989����Karl vonFrisch��1886-1982����������NikolassTinbergen��1907-1988�������ǵ��о��Ȳ�������ѧ��������ҽѧ������ͨ���۲춯����Ϊ�������ۣ��ӽ�ʮ�����͵IJ���ѧ��
vonFrisch�о��۷����Ϊ���ж���������۷������ڵ������ӡ��г��������ʶ��ƫ�������������ý�����Ϊ�����۷�ͨ���赸������Ϣ��һֻ�۷䷢��ʳ��ɻط䳲���������۷�ʳ�����ڵص�Զ���ͽǶȡ�ʳ��Ķ��١�����һ������������Ϊ���̼���ѧ�����۶����Ƿ�������ܷ������Լ�������ʲô�����⡣
Tinbergen�ĸ����1969��ŵ�������ý�������Tinbergen�о�����ı�����Ϊ�����ܵ���Ϊ������������һ����������ֵģ�����һ��ʱ�������ֵ���Ϊ��������ֳ��Ϊ����̵���Ҳ����۲졣Tinbergen��Lorenz���ֶ����п̰嶯������һ�Զ��ڽ������Ҫ��һ���ض���������һЩ�̼������յ��ض�����������Щ�����ɴ̼�����֮Ϊ�ж�������LorenzҲ��������ʵ�飬�������������о�����Ϊ��ӡ����imprinting�������û��ȶ���ʵ�飬�ڸմӵ�������һ��ʱ���ڣ����С��������ƶ�����ֻ��Lorenz���ˣ���ô�Ժ���Щ����LorenzΪĸ�ף����ߵ�������Ǹ���������������������죬������ԥ�������Ǹ���Lorenz���ˡ�ӡ��ֻ�����ض�ʱ�ڲ�������Ϊ�ٽ��ڡ�
1951��Tinbergen����о���Ϊ���ĸ����棬��������ԭ��proximatecauses������������Ϊ����ڸ��巢���������γɣ���������������Ϊ��ԭ�����������ռ�ԭ��ultimatecauses�������������ܣ�����Ϊ������ż��ʳ���
��Ϊ���о����ڶ����кܶ�ͣ���������ij̶ȡ���2009��vonBayern�ȵ����ģ�������ѻ��α�����Сʯͷ�����Լ����������ŵ�ʳ���Ҫ����Ϊѧ�Ľ̿��顶������Ϊѧ��������ƪ���µ�����Ҳ���ࡣ
�������о���ͻ��Ե���Ŵ�ѧ��Ӧ�á��Ŵ�ѧ���ϵ¶���Ħ������1960�������Ҫ�о��ĺ������Ŵ���ԭ������ʾ�����Ⱥ����Ŵ����ɡ���1960���ĩ��ʼ���Ŵ�ѧ��Ϊ��Ҫ�Ĺ��ߣ��ṩ�о����������;����ͨ��ͻ��۲쵽�����쳣���µı��ͣ����ۻ��������Ĺ��ܣ��˽�����ѧ��������������ѧ��LelandHartwell��1939-���о���ĸϸ�����ѵ��Ŵ�ͻ�䣬���۲���ϸ�����ѵĻ��������������������İ�֢��
ͨ���쳣�����˽��������ɣ�ͨ���������ⷢ������ԭ�������Ŵ�ѧ�ĺ���֮һ��
�����ӵĴ��ں�����
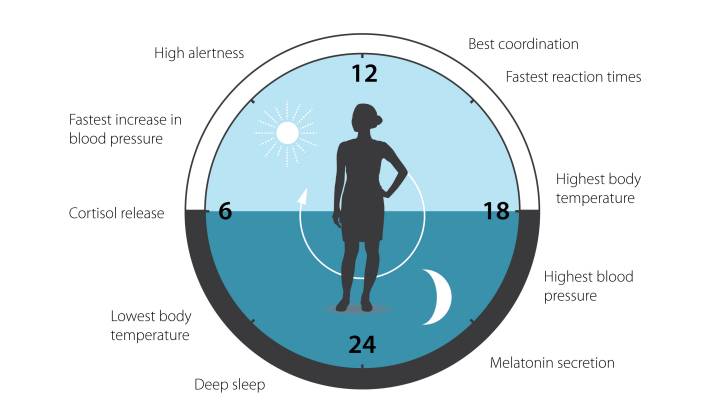
�����������������һ�������ķ��ȡ����ڡ���λ��
�����������ӵ����ڱ��֣������������ڵĶ�ʱ�����������ӱ���������ά�ֵ���������Ϊ���ɷ�������pacemaker������������绷�����أ�zeitgebers�����������Ӷ�ͬ������
�����ж��ֽ��ɣ���ͬ���������Ų�ͬ�Ľ��ɣ�ͬһ����Ҳ�ж��ֽ��ɡ���Щ����ÿ��һ�����ڵĶ��ߡ���Щֲ��ÿ��һ�����ڵij�Ҷ��Ҷ�����ﻹ�и�������������������������������֪�Ľ�������ҹ���ɡ����������Ϥ��˯������ҹ���ɣ��ܶ�������Ϊ������ָ��Ҳ����ҹ���ɡ�
��Ԫ4���ͣ�Androsthnenes֪������������Ҷ���˶�����ҹ���McClung, 2006�����������SantorioSantorio��1561-1636��ǰ��30���¼�Լ����絽������ʳ������й�������ر仯����������ҹ���ɡ�������δ���������Ӱ��Ľ��������������Ľ��ɡ�1729�꣬��������ѧ��JacquesOrtous deMairan��1678-1771���ú��߲������ϣ��۲���ҶƬ�ͻ��ı仯������������ȫ����һ��ʱ�䣬ҶƬ��Ȼ�����кϣ����������⣬������ʱû�����ֲ�����ҹ���������ڵģ�deMairan�� 1729��Szymanski��1918���������Ҳ�о���ֲ��Ľ��ɣ��������ҹ���ɵĿ��Ŵ��ԣ�Darwin andDarwin��1880�������������տ�����Χ������ķ���ת��ͨ��������������Կ������տ���������Ϊ�����ճ�������Ҳ�����ڽ�������������ҹ���ɲ��������ڵ�����ת���ŵ�̫�գ������ճ���Sulzmanet al., 1984����
Aschoff ��Wever��1976���ܽ��˵���ҹ���ɣ��¹���ѧ�����ڶ�ս���µı����У��ƹ�㶨��ʳ��㶨���¶Ⱥ㶨���������ơ�����绷������ȱ��������£�������Ȼ������ҹ���ɣ�ֻ����ʮ�������λ�ͺ�
����֪�����Ӽĵ�ϸ�������壨cynobacteria��ϸ������ϸ�����ˣ��ܶ������������ӵĴ��ڣ�����������Ƿ����е����ϵ����ﶼ����ҹ���ɡ���ҹ�������ڽӽ�24Сʱ������22��25Сʱ�����ڻ�������ȱ��ʱ���Լ���ά�֣����ɵ���λ�����һ�λ�����������⡢���¶ȵȣ���ȷ�����ڻ��������У������ճ����С���һ����Χ�ڣ��������ڲ������¶ȵĸı�����Ҹı䣬���¶Ȳ������ƣ��ⲻͬ��һ��������Ӧ�����¶�ÿ����10�棬ø�Ĵ��������һ����
��ҹ���ɵ�����ѧ����Ŀǰ�Բ��������Ȼ�ڽ��������У������������ת��ƥ�䣬Ҳ�����Խ�ʡ�����������Ч�ʡ������û�����ֻ���ô���أ�����������ҹ���ɣ����������������������������쳣��ϸ����һ�����������������������������ϸ��ռ�˾���������ɴ˿ɼ������Ӷ�������������Ouyanget al., 1998��Woelfle et al.,2004�������Ͻ���о�Ҳ�۲쵽���������̻��ӳ������Ͻ棬���̼���������������뻷���趨��ҹ�������Ǻϵ����Ͻ����ʺϣ�����������ͻ������20Сʱ��ҹ����������ø��ã���������ͻ������28Сʱ��ģ����¸��ã�Doddet al.,2005������Щ�۲죬��ӳ��ֻ�е�����Դ���ڱ���һ��ʱ����������ֲ���������۲쵽��������������ֲ��������ܹ�����Ϊʲô������������Ȼ��֪��Ȼ����֪������Ȼ��
�����������;��
��ô�о������ӣ�
�ܿ������ֲ��ı仯�������ڱ��֣�����֪�������ӵĺ��ģ���������
���Կ����ӵij��롢�����˶��Ĺ켣��������������ָ�������˶��Ļ�еװ�á�
���ڿ�ѧ���õ������о������ӣ���缫��ϸ���۲�������۲쵽�����ض���λSCN���ӽ����Ϻˣ�suprachiasmaticnucleus���ĵ�����ҹ���ڡ��ý���ѧ�о��������˽��ĸ����١���֯����λ���ڸ����������������Ҫ����һ���о��У���������ɹ������Ҫ��Gastonand Menaker��1968��Zimmerman and Menaker��1979�����������ද������ӣ�masterclock������Ϊ��SCN��ȷ�����ӵ�ʵ�������࣬��SCNΪ������������¶���ʧȥ��ҹ���ɣ�Moore andEichler��1972��Stephan and Zucker��1972���������ӻ���ٺ�Ķ���ɻָ���ҹ���ɣ�Lehman etal.,1987�����ڵ��ٺ���ֲ���������SCN���½�������ֲ����Դ�Ķ������һ�������磬�����ֲ����������22Сʱ�Ķ����ô������ֲ�Ķ�������Ҳ��22Сʱ���������ֲ����������20Сʱ�Ķ��������ֲ�Ķ���Ҳ���20Сʱ�����ڣ�Ralphet al., 1990����
������Ŵ�ѧ�о����������ر�����ǰ���Ŵ�ѧ��forwardgenetics���D�D���ɸѡӰ���ض����������ͻ��D�D���������������б���ͨ�����õ�ת�ض������ó��ض�����ķ����Ŵ�ѧ��reversegenetics���������Ŵ�ɸѡ���ŵ㣬����ʱ�����ڶ�������Ϊ�Ŵ�ɸѡ�кܴ��ȱ�㣺�ܶ���Ϊ���²��ǵ����������������ģ����ӵ���Ϊ��Ҫ�кܶ������룬���Ŵ�ɸѡ�Ե���������������Ч���Ը������������Ϊ����Ч���ܲ
���ڹ�Ӭ����Ϊ��JerryHirsch����Ϊ�����õ�������͡�Ҳ�Ͳ���ͨ���Ŵ�ɸѡ�ķ����о���Ϊ��ֻ��ͬʱ�ı�ܶ������д���ѡ�����ƶϡ������ܣ�SeymourBenzer��1921-2007����Ϊ��Ӭ����Ϊ������ͻ�䵥�������ǰ���Ŵ�ѧ�������о���1970����������������������ۣ�Hirsch�������������ڼ�������ѧԺ��ÿһ������д�ųƱ��ܵ��о���α��ѧ��
����
ŦԼ�����IJ�����̫��������ѧ�ұ��ܵĴ�ѧ������רҵ��1945����1953���ν����նɴ�ѧ����ϵ�����о�����ʱ�����ţ��ࣨBenzer,1946����1948������ʼͨ���μ����ڿγ̡�����ʵ������ѧ���1953��ת������ϵ�ν̣��о���������ѧ������������Ҫ�Ĺ������Ŵ�ͻ�����DNA������еı仯��Benzer,1955���������˳���Ӷ��壨Benzer, 1959����

?�������Ӭģ�͡�ͼƬ��Դ��Wikipedia
�ڷ�������ѧ����ŵ�����̶ȵĹ���������Ӣ����ѧ�ҡ�DNA˫������ͬ������FrancisCrick��1916-2004�����Ϸ���̫��ѧ�ҡ�mRNA������SydneyBrenner��1927-������Ϊ����������ѧ����Ҫ�����ѽ��������ѽ������Ժ�ֻ��ϸ�ڸ�����գ�������Ӧ������һ����Ҫ���������Ƕ�ѡ��������ѧ��Crick�����о���ʶ��Brenner������Ҫ�µ�ģʽ�����о�������ѧ������ѡ�������߳棬�������߳���Ŵ�ѧ������һ��ѧ�����߳��о��˷�������ѧ��������ѧ,��Ȼʧȥ��mRNAӦ�û�õ�ŵ��������2002����Ϊ�о��߳������ѧϰ������ѧ�����ù�Ӭ�о�������ѧ��1967�꣬����ת����������ѧԺ������ʼ������Ӭ��Ϊ���Ŵ�ѧ�о����ģ�Benzer��1967����
������1967����2007��һֱ�о���Ӭ����Ϊ����ת���о�������ѧ�ij��ڣ�����ͬ��Ц�����о��Դ����Ĺ�Ӭ���Dz����о����Դ���ë���������ҡ���������ѧ��һ�㲻������������Ѿ��������ͣ����µ������壬���ǿ������ⲻ�𣬲�����������ӣ�����һĻ������Brenner���϶����������������ǵ�ѧ��Ҫô��������Ҫô�������Ҫô������֮�������밲�����ȵ��ˡ�
�������ʮ���У����ܺ�����ѧ�����о��ĺܶ�涼�������磬����ѧϰ���䡢����Ϊ������ͨ�����۷����������������Լ����Ĺ�Ӭģ�͡�ʹ�ȡ��������ļ���ѧ���Ͳ�ʿ���ȫ����������ѧ����Ҫһ֧��
���ܵķ�������ѧ�о��ﵽ��ŵ���̶ȣ������о�������Ҳ���Դﵽŵ���̶ȡ�
͵���Ǵ���֮ĸ
1971���1972�������ƪ���£����Ŵ�ѧ�о������ӡ�Konopka��Benzer��1971������Ӱ���Ӭ�����ӵ�ͻ�䡢Feldman��Wasar��1971������Ӱ��������ֲ����ù�������ӵ�ͻ�䡢Bruce��1972������Ӱ�����������ӵ�ͻ�䡣ͻ����DNA���еı仯��λ���ض������ڲ��������Աߵ�DNA�仯������Ӱ�����Ĺ��ܡ���ͻ����л�����������ǰ���Ŵ�ѧ��ʶ��
����1935���֪����Ӭ��������ɡ���Ϊ��Ӭ���о��߶࣬���о����߶�����������塣��Ȼ�о���Ӭ�����ӵĿ�ѧ��ǰ�����Ҳ����Ҫ�����գ���Ӭ���о��������ӵ��������ƶ����ò���������������壬Ҳ���������κ����
�����ӵĿ���Ϊ��ʱ�����о�����RonaldKonopka��1947-2015���������Rosbash��2015�����ڱ������Ŵ��о���Ϊ��������֮�С�

?Ronald Konopka��1947-2015��
�Ŵ�ɸѡ��ʵ������ǣ����������͵ķ��������±���յ��ض�����IJ�ͬ������ͻ�䣬��ⲻͬͻ���ֵı��͡����嵽��Ӭ�������ӣ���Ӧ���ǽ�����������ӵķ������û�ѧ�ձ���յ���ͬ����ͻ�䣬Ȼ��һֻһֻ����Ӭ���Ա㷢�ָı�����ҹ���ɵ�ͻ���֡�
��ʱû��֪����Ӭ��Լ���������ѧ�����շ����ͻ�䣬ֻ������ֻ��Ӭ���������в���ͻ������ͬ��������Ӧ�üӼ������ſ��ֻܴ���ͻ���һ�Ρ�����Ϊ��Ư�ƣ�һ�������ͻ���֣����ܽ����һֻ��Ӭ��Ӧ�ü�⼸ֻ����ʮֻ����Ӭ�Ƕ����壬ÿ�����������ף�һ���˿��Ա���һ����λ������������������ò�Ҫ����һ����F1����ɸѡ���������F2������������ɸѡ����F2��ֻ���ķ�֮һ��ͬһ����ͻ��Ĵ�������ô��Ҫɸѡ������Ӧһ������������ȫ�������ɸѡ��
����Ӭ��ҹ���ɵı��������ǽ���ֻ��Ӭ�ŵ����Ĺ��У�����ֻ��һֻ��Ӭ�����߶���һ�������С�ܵ����룬���ǹ�Ӭ�߹������룬������ͼ�¼��Ӭ�˶���һ�Ρ��Դ˼���Ӭ��ı仯������ȷʵ����ҹ���ɡ�����ܶ��Ӭ���������Ƚϴ�
Konopka��Benzer��������͵�����Ϸ������ӿ��ٶȡ���һ�ǹ�Ӭ�Ŵ�ѧ���е�attachedX�����Կ��ټ��XȾɫ���ͻ�䡣��������Ǿ�����ɸѡ��ҹ���ɣ���ɸ��һ���ɣ���Ӭ����Ӭ�ܾ��ѳ�Ϊ��̥��������Ϊ�׳棬������Ƥ���ΪӼ��Ӽ������Ϊ�ɳ档
��ҹ������ǹ�Ӭ�ɳ�ÿ�쾭��һ�Ρ���ʮ���ܶ���ʼ��ѭ��������Ӭһ������һ�Ρ���һ��������ǰ�����Ҳ�����硣��ʮ�������ʮ������о���ȷ�����Ϊ�������¶ȵ������������ƣ�Kalmus��1935��1940a��1940b��Pittendrigh��1954�������ǣ�һ��һ�ε�����������ɳ�ÿ��Ļ�����ӣ��Ƿ��йأ���ʱ���������
Konopka��������Ϊɸѡ���������ײ������õ�ֻ��Ӭ���������ҹ����ɼ��Ƚ��������������Ϊɸѡ�������ͺܱ㵱����ѧ�ձ䴦���õ�����ͻ��Ĺ�Ӭ�Konopkaÿ�����粻���ϰ࣬���ʵ���Ұ������Ĺ�Ӭ��������������Ӽ�����絽�������ľ���Ӱ����ɵ�ͻ���֣���Ϊ���ǵ���ʱ�䲻ͬ����������ν��Ұ���͡�����Ӭ�������ķ��������˼�⡢������ɸѡ��
͵�����������Ե�͵��������ʱ���Ǵ���֮ĸ��
�������о���ͻ��
����һ���о�����˵������˼ķ���������ɸѡ����ֻ����ʮ��ֻ��������ֻ��Ӭ����Konopkaֻɸѡ�˲���������Ʒϵ�Ĺ�Ӭ�͵õ���һ��ͻ��Ʒϵ������ɸѡ������ǧ��Ʒϵ�Ĺ�Ӭ�ٵõ�����ͻ��Ʒϵ��
��������ɸ��ͻ��Ʒϵ����Ҫ�����ñ�����˶�����ҹ����֮�������������ҹ��Ƿ�仯���������ȷʵͻ���Ӭ���˶���ҹ�����쳣����Ȥ���ǣ����Ƿ��ֵ�����Ʒϵ��ͻ���Ӭ�ڱ����ϲ�����ͬ��һ��û�н��ɣ�һ�ֽ������ڳ��̼ӿ쵽19Сʱ��һ�ֽ��ɱ�����28Сʱ�����ǽ�һ����ͻ���������е�����ͻ�佻�䣬ȷ������ͻ����Ⱦɫ��ͼ�Ĵ��λ�ã�������ֺܽ������ǽ�һ��������ͻ������䣬���б��ܷ�����˳����⣬��ͼȷ�������Ƿ�ͬһ�������Ǹ��ݵõ��Ľ���Ʋ�����ͻ�������ͬһ������IJ�ͬͻ�䡣���ǰ������������Ϊperiod����дper�������ɵ�Ϊper0��perzero�������ɶ̵�Ϊpers��per short�������ɳ���Ϊperl��perlong����������Ҫ˵�������ǽ��е�˳����Ⲣ���ϸ�ֻ���˷�λ��trans���ļ��û��˳λ��cis���ļ�⣬����ֻ��1971������²�����ȫ�϶�����ͻ���Ƿ�ͬһ������Ȼ��ʱ�Ľ��֧����һ���ܡ�
����ɸѡ�Ĺ�Ӭ�������࣬ȴ�õ���һ����������ֲ�ͬ�����ͻ�䣬����Ե��ر���������
����ɸѡ�ͼ�������ӵķ��������п�����һЩ�������ӹؼ��Ļ����µ�ͻ�������Ӱ�졣ͬһ��������Գ������������ͻ�䣬���������������Ҷ����������Ĺ���Ҳ�ṩ�˺ܺõ�֧�֣��������벻�������������ӵĻ�����Ե���������ͬ�����ͻ����͡��������ͻ��ȷΪһ�����������ͻ�䣬��ôper����ܿ����������ӵĹؼ�����֮һ��
������1940���ĩ��ѧ�������ʦ��1969��ŵ���������¹�����ѧ��תΪ��������ѧ�ҵ�MaxDelbr��ck��1906-1981������ʱҲ�ڼ�������ѧԺ����������˵����������ҵ��������ӵĻ���
Max Delbr��ck: ��I don��t believe it��
Benzer: ��But Max, we found the gene!��
Delbr��ck: ��I don��t believe a word of it.��
��¡per�������ĺô�
1971�꣬��Ϊ����ȱ���������ܵõ�per�����DNA������¡����������DNA������1973��ŷ���������¡��ӬDNA�ļ�����Ҫ�ȵ�1978����˹̹����ѧ��DavidHogness�ͼ�������ѧԺ��TomManiatis���˵Ĺ����ƶ��²��ܽ��С������м����������Ժ�ſ�ʼ�о������ף�Ҳ���׳�Ϊ���磻������������Ϳ�ʼ�о��ǿ��أ����кܴ��Σ�գ�������ȼ�ʮ�꣬���ܿ�����һ��Ҳ�ò��ϡ�
1980����ǻ����¡�ķ������ʱ���������ŶӾ�����¡��Ӭ��per������˷��մ�ѧ��MichaelYoung�������ˣ���Brandeis��ѧ��Jeffrey Hall����������MichaelRosbash����˹��ϣ�������������о���Ӭ��������ʿ�����ʦΪ��¡��ӬDNA������DavidHogness�������DZ��ܵIJ�ʿ����Ӭ����Brandeis�����������ѧר����ͬ����˹��ϣ��������Щ��ѧ�Ҷ����������о����⣬������ͬʱ����Ү³��ѧ��ϣ��������ѧ��SpirosArtavanis-Tsakonas������¡Notch�����������о���Ӭ����Ϊ��fruitless������˹��ϣ�����ý�ĸ�о�mRNA���ӻ�����

?����ֱ�Ϊ�����˶�?�Michael W Young�����ܸ���?������Jeffrey CHall�������˶�?��˹��ϣ��Michael Rosbash�������������2013���ʮ�����ݷ�������ѧ��ҽѧ����
1984�꣬�����˺ͻ����D��˹��ϣ�����ŶӶ��õ�Per�����Ļ�����DNA��Bargiello andYoung��1984��Reddy et al., 1984; Zehring et al.,1984�������DNA���Բ���4.5kb��0.9kb����mRNA�����Կ��ܶ�Ӧ�����������D��˹��ϣ��۲쵽0.9kb֮mRNA�ĺ�������ҹ�仯��Reddyet al., 1984; Zehring et al.,1984������Ϊ��Ӧ��DNAΪper������������Ϊ4.5kb��Ӧ�Ļ�����Per�����������ŶӶ�ȷ��4.5kb��Ӧ����per����Bargielloet al., 1984�� Hamblen et al., 1986; Lorenz et al., 1989����������4.5kbmRNA�Ļ�����DNAת���Ӭ������ʹperͻ���ֹ�Ӭ����ҹ���ɻָ���������һ�����ָı䵼���per����������λ�ɸı��Ӭ��ҹ���ɵ���λ��Ederyet al., 1994����
�������Ŷӵľ��������У�һ������ִ������D��˹��ϣ�����������per����������һ������������Ǿ����Կ�ѧ��ĺô���
�����ѧ�Ҷ�ȷ����per�������������ͻ�����DNA�仯��per0��pers��perl�ֱ�����ǰ��ֹ����������ͬ��λ�ļ���仯��Baylieset al., 1987; Yu et al., 1987a����
�����D��˹��ϣ����й���ѧ����ǿ�����֣�ȱʧper�������ض�һ��������ҹ����ɲ���Ӱ�죬ֻӰ����żʱ��Ӭ����н��ɵĸ質��Yuet al., 1987b��������ʵ������1980�귢�ֹ�Ӭ��żʱ�ij�����н��ɣ�Kyriacou andHall��1980������������飬����������ʽ�����ȶ��Stern, 2014; Kyriacou et al., 2017;Stern et al., 2017����
�����D��˹��ϣ�����ڲ����������о����й���ѧ���У�������ѧ���Ʊ�ҵ����ǿ������ҽѧԺ��������Liu et al., 1988,1992���������Ļ���ʯ ��Huang et al., 1993�����人��ѧ���������Zeng et al., 1996����
per�Ĺ��ܣ������Ļ���
�õ��������˹��衣
���ǣ��õ�����һ�����ܹ�����֪������������������������ֻ�ǹ��ߣ�������������ͳ������������һЩ�ܸ߳бȽ����ĿƼ��������յ��³Ʋ��������������ǽ�ʾ������ء����������İ��ػ�ԶԶ�����������������Ҳû���Զ��������ء�
ͬ�����õ���per�����DNA������֪��������������ĵ����ʵĹ��ܣ�Ҳ�����Ƶ��������ӵĻ�������ʱ���õ������ܳ�ʱ�䶼���������书�ܣ�Ҳ�Ͳ�����������ѧ���̻������༲���Ļ��������磬�����ᣨLesch-Nyhan���ۺ����������ԲУ��Ǻ�����ļ�����������Ѿ�֪�����꣬����HGPRT���λ������������ת��ø�����Ǻ����л��һ��ø����֪������֪�������ʶ����ܸ�������Ϊʲô���˻��ԲУ�Ҳ���ܸ��������вк��������������Ǿ�������������Բе�����ѧԭ��
Ҫ�Ƴ�������뵰���ʵĹ��ܣ������Ԥ�Ƶİ���������������ij�ֵ������������ԣ���ô���ܾͿ������ƣ��Ӷ������ṩ�о������ֵ㡣�����鶼����per�������в����������ظ��ģ�������ʵ���һ���Ϊ��������С������У�Shinet al., 1985�����������ֲ�����ˡ�
1986�꣬���������鶼�������³�per�������ǵ����ǣ�Jackson et al., 1986; Reddy et al.,1986����������������鶼���ˣ������Ŀ��������һ���������¡�
1987�꣬�����˿������밮��˹̹ҽѧԺ��David Sprayʵ���Һ�������һ���������£�Bargiello et al.,1987������PerӰ����ϸ���佻������ʾper�������϶������صĹ��ܣ������϶���ף�����һ����ܿ����ǴӲ²�per�ǵ����ǨD�D������Ĥ���רD�D���Կ����Ƿ�϶����DZ��ʶ��ɡ�����һ��������ظ���1992�������˺�Spray�����ŶӲ��ò���Ǹ���ջ�1987��������Ŀ�ġ���Ȼ�����£�Saezet al., 1992���������D��˹��ϣ����1993��Ҳ��������˵��Per������ϸ�������ӣ�Flint et al.,1993����
��1984��1987���ڼ��Ҿ����Ĺ����У����������鶼�������Σ����Էֱ����һ�Σ�ͬʱ����һ�Ρ�ͬһ���о��������εıȽ��٣���һ���Ĵ��������Ρ�
per mRNA���о�����ϸ���ǻ���
��¡����˻����ٿ�����ı�������Ȼ�����졣
��������ѧ�ġ����ķ���DNA�DRNA�D�����ʡ�DNA�����Ŵ���Ϣ�����Ը��ƺ��Ŵ�����DNAָ���£��Ŵ���Ϣת¼Ϊ��ʹ���Ǻ��ᣨmRNA������mRNAָ���ºϳɵ����ʣ�������������ֹ��ܡ���ͬ����ת¼������ͬmRNA���õ���ͬ�����ʣ�������ͬ���ܡ�
�����������Լ��mRNA�ı��Ҳ���Լ�⵰���ʵı��һ����˵��һ������Ĵ�ϸ������ͬ����DNA�������в�ͬ��mRNA�͵����ʡ������D��˹��ϣ�顢�������鶼���per��mRNA�͵����ʱ�����ʲôʱ��Ϳռ䣨Jameset al., 1986��Saez and Young, 1988��Liu et al., 1988��Siwicki et al.,1988��Zerr et al., 1990��Liu et al.,1992�������з��˺ܴ�ľ�����per�����ʵĿ��壬��ΪmRNAֻ�ܿ��������������ϸ��������������Щ�����ϸ�������ǹ�Ӭ�������ӡ���mRNA���ܿ�������ﵰ�������ڵ���ϸ����λ�����������ʶ���ʶ�ȷ��per��������ϸ���ڵ�λ�á�per�����ʺ���ȿ��Դ�����ϸ���ˡ�Ҳ���Դ�����ϸ���ʣ�Saezand Young��1988��Siwicki et al.,1988������1992�꣬���������²�ȷ��per�����ʴ�����ϸ���ˣ�Liu et al., 1992����
��Ȼper���������ҹ���ɣ���ôper�Ļ�����mRNA���ߵ����ʣ��Ƿ�Ҳ��ʱ����أ�������ҹ���1988�꣬�����D��˹��ϣ�����per�ĵ���������ҹ�仯��Siwicki et al.,1988����per��mRNA�Ƿ�����ҹ�仯�������ϣ��о�RNA�仯���о����������ף���Ϊ�������ڻ�úõĿ��壬��ֻҪ�õ������DNA�ͺܿ����ͨ����ͨ�ķ�������ѧ�����о�mRNA�ı�������δ��per��mRNA��ҹ�仯��һ����������ʵ����˼������á�����ϸ��һ��������żȻ�ģ���˵������permRNA���ˣ��Ǵӹ�Ӭȫ�����RNA��ֻ����ҹʱ�䡢�������岿λ��������ͻ�Ƶ�1990�꣬�Ǵӹ�Ӭͷ����ȡRNA�������������֣��縹��������RNA�ڸ���ͷ��mRNA��
Ϊʲô��Ҫר���о�ͷ���per����?������ҹ���ɵĹؼ����ִ���������ʲô��λ�йء��ڵõ�per����ͻ���Handler��Konopkaר���ڲ�ͬͻ���͵Ĺ�Ӭ֮����оֲ�ϸ������ֲ���Ա�ȷ���ĸ���λ����������á���pers��Ӭȡ������֯����ֲ��per0��Ӭ��ǻ�У�����per0�����pers����ʾ��ȷ�����ɣ�HandlerandKonopka��1979����1983�꣬�뿪��������ѧԺ��Clarkson��ѧ��Konopka�������Ŵ�Ƕ������ʵ�飬�ù�Ӭ���������ϸ����ͻ���per����pers���������ಿ����Ȼ������������֣�����Ժ�pers����Ӭ�������Ӿͱ�̣�Konopkaet al., 1980�������ԣ���ֲʵ���Ƕ����ʵ���֧��per���������п���ȫ������ҹ���ɡ�
1990�꣬�����D��˹��ϣ��IJ�ʿ��PaulHardin����ÿ��Сʱȡ��Ӭ��ͷ�õ�mRNA���ټ��per��mRNA������������ҹ�仯��Hardin��Hall��Rosbash��1990������pers�У�per��mRNA��ҹ����Ҳ���̡����������ģ�ͣ�per�Ļ���ת¼per��mRNA���������per�����ʵĹ��̴��ڸ�������per��mRNA���ʲ�������Ӱ��per����������ת¼�����ǵ�ʱ�����ų�per������ҹ������һ��Ϊ֮����Ϊ��ͨ�������ӵķ���������per����ת¼��������Ҫ��Occam�굶ԭ�����ȼ����Ŀ����ԡ���һ�����ṩ�������ӻ��Ƶ�����ͻ�ƣ�������Ϊֹ�Ա����ϵġ�ת¼�D���븺������·��trans***ion-translationfeedback loop, TTFL�����Ļ�������˹��ϣ���������һģ�͵IJ�ʿ��Hardin��
�����һ������ȷ����ôper�����ʾ��ǻ����ת¼�������ӡ�������ת¼���ӣ�1980����кܶ��չ���Ӷ�����Ϊ�о�per�������ӵĻ����ṩ�ܺõĽ����
���ã������D��˹��ϣ���Zerr���ÿ��������per�����ʵı���ڲ�������Ҳ����ҹ���ɣ����ֽ��ɵij���Ҳ��per���������ڣ���Ϊ�����������̵Ĺ�Ӭͻ����pers�У�per�ĵ�������ҹ����Ҳ���̣�Zerret al.,1990����mRNA����ҹ�����뵰������ҹ����֮����һ���ļ����Ҳ������λ��ͬ������֮��֪������per�����ʺ����仯����������ϸ����λ�仯����ҹ�����У�per�����ʽ���ϸ���˵�ʱ������ҹ���ڱ仯��Curtinet al., 1995����
����ת¼����
1990�꣬�о���Ӭ�����ӵ�Hardin�����per�����ʿ��ܵ��ڻ���ת¼�����ݵ��Ǽ��֤�ݡ�
1991���1992�꣬����;�����о������˼�����per������������ĵ����ʡ��о���Ӭ��̥������Crewsʵ���ҷ���һ������singleminded��sim������ȱʧ���¹�Ӭ��̥�������ߵ���ϸ��ȱ����Nambu et al.,1990�����о������°����Hoffman�Ⱥ�Burbach�ȣ�����һ�����°������Ӣ(dioxin)���õ�����AhR��Hoffman et al., 1991��Burbach, Poland, Bradfield, 1992����per��AhR��sim���������ʶ����Ƶ������ΪPAS����
��ΪAhRȷ����ת¼���ӣ�������ר�ŵ���ת¼������bHLH��������ʾper��SIMҲ��ת¼�������ӣ���perȱAhR���е�ת¼��������ֻ��PAS����1993�꣬��˹��ϣʵ���ҵĻ���ʯ֤����PAS�����ǵ����ʨD���������������PAS��PAS����ż�ۻ���Huang,Edery andRosbash��1993������һ���о����������bHLH��PAS����ĵ����ʿ�����ת¼�������ӣ���ֻ��PAS��ȱ��bHLH��per�����ʲ��ܼ���ת¼�����������ܹ�����ת¼��ͬʱ��bHLH��PAS����ĵ����ʣ�Lindebro,Poellinger and Whitelaw��1995����
Hardin�Լ�����ʵ���Һ�֤��per�����ʵ���per����ת¼��������per�����ڵ����ʱ�������֮���Σ���������ѧ��Ϊ5���ˣ���DNA���У�Hao,Allen and Hardin, 1997�����Ժ��и���֤�ݱ�����per����ת¼���ڣ��������������йء�
��������Ӭ�����ӵĻ���
Ӱ�������Ӳ�����ֻ��һ������
1971�귢�ֵ�һ��Ӱ���Ӭ�����ӵĻ���per��
1983�꣬Jackson������Ӱ���Ӭ�����ӵ���������Jackson��1983�����������ա�
1990�꣬�����D��˹��ϣ�鷢�����¡��Ĺ�Ӭ��ҹ���ɻ����ӡ���Clock����Dushay et al.,1990��������������1992�귢���ⲻ���µĻ������Ϊper�������һͻ����ѣ�Dushay et al.,1992��������Ŭ����Ϊһ����ˮ��
1991��Konopka �ȷ����ڶ���Ӱ���Ӭ�����ӵĻ���Andante ��Konopka, Smith andOrr��1991����ע����һ������˲��࣬��2003�����ű�Tufts��ѧ��Rob Jacksonʵ��������¡��Akten etal., 2003����
1991����˷��մ�ѧ������ʵ���ҵ�Sehgal���ڻ����ϱ�������ɸѡӰ���Ӭ�����ӵ��»���Sehgal et al.,1991����
1994�꣬������ʵ���ұ������Ƿ�����timeless������ʱ�䡱�����Tim�������Seghal��ͨ��ɸѡ7ǧ���ͻ�����ҵ�������Ӱ���Ӭ�����ӵ��»���Seghalet al.,1994������������ת���Ӳ����յ�ͻ�䣬������¡��������ļ��Ҳ��������û���ɺ�ʵȷʵӰ������ҹ���ɡ����ǻ�����tim��ͻ��ҲӰ��per��RNA��ҹ�仯��������per������ء�������ʵ���ҵ��о���LeslieVosshall����TimӰ��Per�����ʳ���ϸ���ˣ�Vosshall et al., 1994����
������ʵ���Һܿ��¡��tim����Myers et al.,1995������ȻԤ�Ƶ�tim���������п�����������ô���õĵ����ʣ�����Ϊ�����Խ��per����ô����Ҳ��ת¼���ӡ���˹��ϣʵ���ҵ��о������������tim��per���������ʵ����������ҹ���ɣ�������Ե���tim�����ʵ��ȶ��ԣ��Ӷ��ṩ�˹�������ӵĵ��ڵķ��ӻ�����Zenget al., 1996����
��������������ҹ���ɵĻ���
���������ϸ����ֲ�ﵽ���������ڵĶ�����н��ɡ�����ҹ���ɡ������ڶ�������������ӣ��Ƿ���ͬ���ķ��ӣ��Ƿ���ͬ���Ļ�����
�ֲ����ù��neurospora���������ӻ���Frequency��frq����Ƶ�ʡ�����1989�걻��¡����ʱ��Ϊ����per�в����������ƣ�McClung, Fox,Dunlap��1989�����������ֲ����ơ�
������Щϸ�������ӵĻ���Ҳ�ױ���¡���磬Liu et al., 1995����
1995�꣬����ֲ������Ͻ棩�����ӵĻ���¡��Millar et al., 1995����
�ܶ��˹��ĵ��ǣ�������������ӣ���ʲô����
ǧ����ʼ���������鶯��������ӻ���
���ڲ��鶯�������ӵĵ�һ������tau��ͻ������1988�걻żȻ���֣�Ralph andMenaker��1988�����������¡��Ҫ�ȵ�2000�꣨Lowrey et al.,2000����������һ������ø��CKI�ţ���
�ڿ�¡�˹�Ӭper�����1984����1997�꣬�к�ǿ�ҵ�ϣ���ҵ������������ӻ�������Ӧ������per�Ļ�����һ;���Ѿ���ţ����֮�������������ɹ������ҵ��˲ϵ�per����Reppertet al., 1994���������������Ҳ������per��
ʱ������Ӣ�ۣ��ҵ������������ӻ����Ӣ�ۡ�
������֥�Ӹ籱��������ѧ�����������ѧ�Ҹ��ž��������������ķ����������������Ŵ�ɸѡ��
��Ӭ��С���������ڶ̣������ұ��ˡ���Ҫ��ܶࡣ���ͨ��ɸѡ�һ�����ô�������Ӭһ������������Ҫ���Ǿ��ѡ�Ϊ����һ����Χʡʱ��;��ѣ�����ʵ���Ҳ�ɸѡ������ͻ���֣���ɸ�Ӻ��塣����ɸ�Ӻ�����Ȼʡ�˹���������Ҳ������ͻ�����ʡ���ʹ������Ҳ��Ҫ����ʮ��ֻ���������ɸһ�顣�и�Ц����������ʵ�飬���ưѺ����ù�������
1994�꣬����������ûһ��ʵ������Ϊ�Լ����㹻�ľ��ѡ��ܿ��������˾��ѻ��Ҳ���Ӱ�����������ӵĻ�������û�˸�����
���Ŵ���ʵ�����Ƿ���ͼǿ��������������?
���ǵ��������������������ǵ�ʱһ��ֻ�о���304ֻС��25ֻ����Ӱ�������ӵ�ͻ���壬���ǴӶ�������Ӱ�����������ӵĻ�����������Ϊ���ӡ���Clock����Vitaternaet al.,1994�����������������ӵ�������23.7Сʱ��Clock�Ӻϵ�ͻ������ҹ����Ϊ24.8���仯��ν���Ҫ�ɿ��ļ����ܷ��֡����Ӻ������С���ɿ��ı仯����ʵ���Һ�����ͨ������С������Clock����ͻ��Ĵ����壬����ͺ�ǿ����ȫɥʧ���ɡ�
1997�꣬����ʵ�����ڡ�ϸ������־������ƪ���ģ��������ǿ�¡������Clock����King et al., 1997��Antochet el., 1997����
����Clock�����ʲ������뵰��������õ�PAS��������ֱ�ӽ��DNA��bHLH����King et al.,1997�����Ӷ���������������ε��ڻ���ת¼��Clock������Ҳ������SCN�����ǻ������ˡ��������桢�ܡ���Ҳ��Clock����Kinget al., 1997����
��Ӭ�벸�����Ƿ������Ƶķ��ӻ���?
�����Ӭ��per����ֻ���ڹ�Ӭ�����������ã���ô��������ޡ�����ҵ��ߵȶ����Per����˿����о��ߵȶ��������ӵķ��ӻ�������ô����ͽϴ�
��Ӭ��per������1984�걻��¡�������ˣ�����ʡ��ѧ��˹��У����StevenReppert����ͼ�ڸߵȶ������ҵ�Per��������һ����˵��һ�������ڵ͵ȶ����з��ֺ��ڸߵȶ����бȽ������ҵ����ж��ַ�������ͨ��DNA�����������ҵ����Ҳ����м��ֿ��ܣ�Per�ڸߵȶ��ﲻ���ڣ������Ӭ�������ӿ��ܸߵȶ���������Ӳ�ͬ��Ҳ�����ǹ�Ӭ�ߵȶ��������Ӵ������Ƶ������з��Ӳ�ͬ�����Ӭ��per���ߵȶ��ﲻ�ã�����һ�������о������ϵ����⣬�ڸߵȶ������Per����δ���ҵ�����1984�굽1997���Ŭ�����Ҳ������鶯���per���������£��ܶ��˻��ĵ���Ϊ���²�������Per��
1997�꣬����ʵ����������ţ��ҵ�������˵�Per ��Tei et al., 1997��Sun et al.,1997�����������˿�ѧ�����о����������ʱ����һ��������per�����������ԣ�Sun et al.,1997�����Ƚ����Ǻ�֪������ǰû�ҵ���ԭ���Dz������Per���Ӭ��per��������������������Բ��ܸߣ����ֲ��ܸߡ���ǰ������ȥ�Һ����ҵ������þֲ��������ڵ�ʱ����֪���Ķ��DZ��صġ�
�������Per���������� ��Per1��Per2��Per3��Shearman et al.,1997����Per���������SCN��Shearman et al.,1997�������������ҹ���ɱ仯���仯����һ������Clock����ĵ��ڣ�Jin et al., 1999����
1998�꣬�����D��˹��ϣ��ͨ���Ŵ�ɸѡ��Ҳ�ڹ�Ӭ�ҵ�Jrk�����־��ǹ�Ӭ��clock����Allada et al.,1998������Ȥ���ǣ�SteveKayʵ���ҷ���clock�����Լ������per����������Ƭ��E-box������ӫ���أ�luciferase���������ı����һ�����ܱ�per���������ƣ��Ӷ��Ϻõ�ʵ�ָ�������·�ıպϣ�Darlingtonet al., 1998����
�����ڹ�Ӭ���ֵ�per���������ڲ������ҵ��ˣ��������������ֵĻ���ClockҲ�ڹ�Ӭ�з����ˣ������浽�����࣬�����ӵĻ������߶ȱ��ء�����Ȼ�ǻ���ˮƽ������ʾ����Ҳ���ء�
���о���˵�����й�Ӭ�����ӵĿ�ѧ���ǾͲ����ٵ������Լ�����ȷʵ������̽�ֺͽ�ʾ������ͨ���ձ������
��Ӭ�Ŵ�ɸѡ���������о�����Ҫ��
�������ӵĻ����о������У���Ӭһ�����ȣ���ѧ�Ҵ��з�����per��Konopka and Benzer��1971����andante��Konopka, Smith and Orr��1991����tim��Sehgal et al., 1991, 1994����
�ù�Ӭ�о������ӣ����ٶȡ��۸������϶����ڲ����ࡣ��ϸ���������ֲ���о������ӣ�Ҳ��������Щ���ƣ������ǵ������ӻ����붯��IJ�ͬ���о����Dz��ܽ�ʾ������������ӻ�����
��Ȼ���ڴӸ�������õ���֮ǰ��������Ԥ�������ӵĻ������صķ�Χ�����º�����ܽ��Ӭ�����������о�����Ҫ�ԡ�
�������е͵�������о��������ڸߵ������Щ��ͨ����Щ��ͬ�����磺ϸ�����ڵ�ԭ���ӽ�ĸ���˶����أ����Ա�ȷ���Ļ����ӹ�Ӭ�����ಢ�����ء���Ȼ�о��͵��������е�����ѧԭ��Ҳ�п�ѧ���壬����ƫ�ڹ�ע�Ըߵ��������õķ��Ӻ��ɡ�
�ڷ���per��andante��tim�Ȼ���֮��ѧ���ǻ�����ù�Ӭ����ɸѡӰ�������ӵĻ���
����ʵ���ҽ�����ӫ���ؽ���per����λ�㣬��ø���Լ�ӱ���per��ת¼�����Brandes et al.,1996��Stanewsky et al., 1997�� Plautz et al.,1997����ɸѡ��5137��Ӱ��ӫ���ر���Ĺ�Ӭͻ���֣�����chryptochrome��cry����������Ӭ�����ӣ�Stanewsky et al.,1998�����ù�Ӭ���Ŵ��о�����cry���������ڽ鵼���Ĺ�����������ڵ������ӣ�Stanewsky et al.,1998������������û����绷��Ӱ�������£�����������ת������绷�������£����������������ϡ���Ȼ�������������¶ȡ���ʳҲ��Ӱ�������ӣ����ĹȻ�ǵ��������ӵĹؼ�����֮һ��cry���ܵķ��ֽ����һ����Ҫ�����⡣����Ҳ���Կ������Ŵ�ɸѡ����Ϊ���ҵ���������ʱ���ܴӻ������ĵ������Ʋ�����ӻ��������˽����������ѧ�����о��ĺ��ġ�cry������mRNA����Ҳ����ҹ���ɣ���Ҳ����per��tim�������Ĺ�ϵ��Emeryet al., 1998��Stanewsky et al., 1998����
��Ӭ��4��Ⱦɫ�塣��˹��ϣ�D��������ɸѡ������Ⱦɫ�壬�û�ѧ�ձ�õ��Ŵ�ͻ���֣�ɸѡ��6ǧ�����Ժ���jrk����Allada etal.,1998����������ο���ǣ���Ӭ��jrkԭ������С��Clock�����ͬԴ�������ҵ���cycle����Ҳ��С������ƻ���Bmal��Rutilaet al., 1998�����ٶ�֤ʵ�����浽��������ͬ���Ļ�����������ӡ�
������ʵ�����ڹ�Ӭ�ĵ�2�͵�3��Ⱦɫ��ɸѡ�˶���1��5ǧ��ͻ�䣬��������������Ϊdoubletime�ģ�����ʱ�����������������ҵ���ø�ţ�CKI�ţ���Priceet al.,1998���������Ե���per�����ʵ��ȶ��ԡ�����ʵ������2000���¡�˽�����tau������ԭ��Ҳ��CKI�ţ�Lowrey etal., 2000������ʼ�о�����ø�������ữ�����ữ���ڵ������ȶ��ԡ�
��2001�꣬��֪��������ڹ�Ӭ��С���������ӣ�������Ӭ��һ��per��С�������Per����Ӭ��һ��cry��С�������Cry����Ӭ��һ��jrk/clock��С�������Clock����Clock��Npas2������Ӭ��cycle��С���Bmal1����Ӭ��doubletime��С���CKI�ĺ�CKI�Ż���Reppertand Weaver, 2001����
ɸѡ��������ػ��������;��
����ȫ��������ӻ���������£����Ŵ�ɸѡ�����ӣ��Ǻ�ǿ�ķ������ӷ����ϣ���֮Ϊ��äɸ����δ�����ɡ�äɸ���ŵ�������������֪����������ͨ���Ŵ��õ�ͻ�䡢DNA��¡�õ����ӡ����ӵ���������������о����ٽ�ʾ���������ṩ��ʾ�����Ŀ����ԡ�����һЩ����ν�ġ��Ŵ�ѧ�����˾�η����������theawesome power of genetics������һ;�������ڶ����Ҫ������о��������������ڵ��о�����֮һ������Ψһ��
������������ʱ����������ͻ�Ƶ�ʱ����ȻҲ����ٶ��Ŵ�ѧ��Ψһ;����
���������о��У�������֪��Per��mRNA��������ҹ���ɣ�Per��������Tim������������ã�����ø���������ӣ���ô����ͨ����Щ��������������Abruzzi et al.,2017������������չ�����������ѧ��RNAi��RNA���ţ�������֮��Ҳ��Ȼ�������¼����о������⡣
��ʵ�ϣ�����1995�꣬����ҽѧԺWeitzʵ���ҵ�Gekakis��ͨ��Per������Ѱ����Perֱ������õĵ����ʣ��ҵ�Tim�����ʣ�����Tim��Per����������ֱ�ӽ�ϣ�Gekakiset al.,1995���������ý�ĸ˫�ӽ��õ�����һ������ͬ��������ʵ�����ù�Ӭ�Ŵ�ɸѡ��;������ͬ��������ʱ������ʥ���Ǹ���ݴ�ѧ��UCSD��SteveKayʵ��������ʿ����Ŷ����״���RNAiɸѡ�����ӵĻ��������õ�ʱ��֪����ϸ���������ӣ�������������������ϸ����Per�������ӫ���ػ���ͨ��ӫ���ػ���ı���������ʾ�����ӣ��Ŷ�������ҵ�Ӱ�����������ӵĻ���Zhanget al., 2009������Ȼ�û����Ǻ���ɸѡ����ģ���������������ϸ���������ˡ�
��1971��Ĺ�Ӭɸѡ��2009���˵�ɸѡ���о���ʱ���Ⱥܴ�����û����ȫ����Ӭ��ɸѡ��û��ɸ�꣬����ÿ�ζ��о��ޣ����鶯���ɸѡ��û���ꡣ����Ժ��и��ࡢ���õ�;���ͷ������������к��Ѿ��ù�����δ���ʹ�õķ������ٷ���Ӱ�������ӵĻ��������˾��ȡ�
���ӻ�����ϸ������
���ֻ����Ȼ��Ҫ��
������ѧ�����о���Ŀ���Dz���������
�ۺ϶�����о���������Կ�����Per���ڻ���ת¼���Ƿ��ӻ�����ͻ�ơ������֮��Clock��Cycle��������ɸ����壬ֱ�Ӵ̼�һЩ���ΰл����ת¼����Щ�л����һ�����������ӡ���Per��Tim������������Clock��Cycle�����á�Per�����Tim����������Clock��Cycle�İл��Ӷ����ת¼��������Per�����ʵ��ȶ��Ա����ữ�����ڣ�Ҳ�ͱ�doubletime��CK1�ĺͦŵȵ���ø��ȥ�����ữ�ĵ�������ø�����ڣ�Wijnenand Young��2006��Benito et al.,2007���������ǻ����ε���Per��Clock�����ʡ�Cry�鵼�����Tim/Per���ȶ��ԡ���Clock/Cycle��Per/Tim�������Ҫת¼��֮�⣬��������ת¼������������������pdp1��vri��RORa��REV-ERB������ת¼���ء�
Per��Tim�����ʲ������ȶ��Ե����⣬���һ�����ϸ����λ���⣺��ϸ����������ϸ���ˡ���Ϊת¼�������ӣ�����ֻ���ڽ���ϸ���˲��ܼ���������ڵĻ��������ܵ��ڻ���ת¼��Siepkaet al., 2007��Dubowy andSehgal��2017����Per��Tim��mRNA�ڰ��������ߣ������������߷塣���������ʵĺϳ�������mRNA��Сʱ�����ۣ�����ϸ��������ҹ����ϸ���ˡ�Tim�����ʼ���ϸ�������ȶ�Per�����ʣ�Ҳ�ٽ�Per����ϸ���ˣ�Meyer,Saez and Young��2006����
��Ȥ���ǣ���Ȼϸ�����������ֲ��ֱ�������������������ӣ����Ķ�����������Щ����������ɲ�ͬ�ķ�����֣�Ҳ��ͨ����������������ʵ�֡����Ĺ����������ͨ��һ����ʽӰ�������ӵ�����Ҳ����ת¼���ء������ữ���ڵȣ�������Wijnenand Young��2006������ֲ��������ӻ�����ϸ������ϸ���ʶ�λ�ĵ��ڣ�Wang andTobin��1998��Mas��2008������ˣ���ϸ�����˲�ͬ����������Ӵ���ԭ���Ĺ�ͬ�ԡ�
��ϸ�����棬Ŀǰ��Ϊ���������ھ������ϸ������������ϸ�������������ӣ�Emery et al., 1997��Giebultowiczand Hege��1997��Plautz et al.,1997������Ӭ�ij�����º���Ȼ���Թ۲쵽���е�������Ӱ�죬�˵ĸ���ϸ����������Ҳ���ֳ���л�ϵĽ��ɡ�������Ϊ�����ӷ�Ϊ��������Ӻ����ܵ��ӡ�
�ڹ�Ӭ���ڣ��Ʋ��Լ150������Per��Tim�������ϸ��������ӣ�masterclock�����ڲ�������SCN������ӡ�С���SCN��Լ�����ϸ�����˵�SCNԼ����ϸ���������ӿ���������ת�������ӿ���ȫ���������ӣ��ر���ͬ���������ӡ�
Konopka��Benzer��1971������¾�ָ����ͻ����������õĽ��ʲ�λ����Ҫ��Ҳ����˵����ܹ��ҵ���Щϸ�����Ϳ����ҵ��������ӡ�һ���Ʋ��Ӭ�Ժ�Լ25�����ϸ����������150��ϸ�����������ӵ��Ʋ�������Per��Tim�Ĺ��ܺͻ�����ﲿλ��Konopkaand Benzer,1971��Vosshall and Young, 1995��Kaneko��Helfrich-Forster,Hall��1997��Panda��Hogenesch and Kay��2002��Helfrich-Forster,2005�������ǰ������������ԪDN1��DN2��DN3�������ԪLPNs������ƫ����ԪLNd������ƫ��С��ԪsLNvs������ƫ������ԪlLNvs������lLNvs��Ԫȫ����PDF���ģ���5��sLNvs��4����PDF��Nitabachand Taghert��2008����PDF��������Ҳ����Ҫ���ã�Renn et al.,1999������Щϸ���Ĺ���Ҳ��һ������PDF��sLNvsϸ��Ϊ���ƹ�Ӭ���ϻ���糿ϸ������LNd�Ͳ���PDF��sLNvsϸ��Ϊ���ƹ�Ӭ������İ���ϸ����
���鶯��������ڲ�Ҳ�зֹ���Э����SCN����ϸ�������״̬�£��ֱ��н��ɵ���ͬ������������һ��ʱ����ͬ���������ĵȷ��Ӳ��룬�������硣
�˵���������ػ���
��Ȼ�����Ӷ��˺���Ҫ�������ǽ��������˵����������⡣�Ŵ��ͻ����������ͻ���Ե����ã��������Ӭ���ơ�
��8753�Գ���˫��̥���о����棬�˵���ҹ�������ǿ���Ŵ��ԣ�Koskenvuo et al., 2007����
2001�꣬�ɽ�ɽ���ݴ�ѧ��UCSF����Pt��cek��Fu��������ʵ���ҷ��֣��˵�Per2����hPer2�����˵���������ء������о���һ���Ŵ��Ե���λ�仯������˵���λ��죨������˯����λ��ǰ�ۺ�����FASPS����ÿ������4����ѣ�����7���Ҫ˯����λ��һ������ǰ4Сʱ��Pt��cek��Fu�Դ˼�ϵ���Ŵ��������֣������з���hPer2������ض�λ�㣨��Ӧ���䵰�������е�662λ�����ᣩ�仯�ͻ���λ��ǰ�IJ����������λ���������ˣ���û����λ��ǰ����Tohet al.,2001����662λ����Ӧ�İ�������˿���ᣨ���S�������ǵ�ʱ��ΪS662����CKI�������ữ��2005����Pt��cek��Fuʵ���ҹ��������ν������ݴ�ѧ����謷���CKI�Ļ���ͻ��Ҳ�����˺�С������λ��ǰ��Xuet al., 2005����
����и�������ֲ����˵������ӣ�He et al.,2009����������ʵ������2017�귢���˵�Cry1����ͻ����Ե�����������λ�Ƴ٣���˯���𣩣�Patke et al.,2017����
��������������������
�������Ŵ��о�ȷʵ����������Ҳ��������
�ڿ�����ǰ;ʱĬĬ̽������һ��С�����ڳ��ڸ��ţ������ѳ�������������⣬�����������Ҳʱ�����á�
�ֿ��ƺ�������Ҫ��ϸ���ɼ�������Ҫ��������ǰ��û����������û�����ΰ�˲����ŵĹ�����
Konopka��Benzer����ǧֻ��Ӭ��ɸ����ֻͻ���Ӭ����Ȼ��ͬһ�����������Ӱ����ȫ��һ����ͻ�䡣�Ŵ�ѧ�о�����ʷ�Ͽ���ֻ����һ��С��ģɸѡ�г���һ��������������ܸı�����������ѧ��ʷ�ϣ������Ҳ�Ǿ����У�˵������ѧʷ��Ҳ�Ƿ�ë��ǡ����û��������ͻ�䣬���ܻ���Щ�˻�Ƚ���ԥ�������ܷ����Ŵ������о�����Ϊ��������һЩ�ڹ�Ӭ������ɸѡ�������б��ͣ�����һ����ӳ�����Ӻ��Ļ����Ļ������ܷ�ɿ���ֱ�ӵذ����Ŵ�ѧ���о��������⣬��һ���dz���Ҫ�����⡣��Ӭ�����ӵı��ͣ���ʮ���Ľ��쿴���dz��ɿ��������������������������ʣ��ӹ�Ӭ����˯�ߣ��Ƿ�����˯�ߣ��ܶ��ˣ������о���Ӭ�Ŀ�ѧ�ң������������ػش���һ���⡣
����ʵ�����������ɸѡ�����У���25ֻ�������Ӱ�������ӵ�ͻ�䣬Ҳ�кܴ�����������û����������������Ҫ���ܶ��ʱ�䣬����������о��ܷ�Ǯ��û���㹻�����������������ɡ�һ����Ԥ�Ƶ��ɱ����Ͳ������������о���
�������ǿ����º���������ܽ��Ժ���ʵ�����ʶ����ʵ����ֻҪ���㹻��ɸѡ������ɸ��ǧֻ��Ӭ������ɸ����ֻ��Ӭ������һ�����ҵ������ӵ�ͻ�䡣��Ȼ����õ�һ����������ֲ�ͬ�����ͻ�䣬��ͬ�������ͻ�ƵĿ����ԡ����ɸѡ��Ӭ������ͻ���ֵĶ��ɸѡ��ֻ�����αȽϵ�ù��һ�����ظ�������ǰ�Ļ���һ�θ���ˣ��������ҵ����������µĻ���
����ʵ����������ɸѡ������ͻ�����Ҳ�����������º���������о��ġ����ѹ���ɸѡ������ǧֻ��Ӧ���ܵõ�Ӱ�������ӵĻ�����ǧֻ����ľ��ѣ�����ֻ�и���ʵ���Ҳ��С�����һ������ܽᲢ��ͽ�ͣ�ԭ����������ҽѧ���ġ������ձ���������˯��ҽѧ�о�����MasashiYanagisawa�����Ŷӣ����ýϴ�ר��ɸѡС��˯�ߵ�ͻ���֣��ҵ�Ӱ��С��˯�ߵĻ���Funato et al.,2016����
Konopka���о�����������������ѧ��ǰ����������������˻���ȴ�൱���á�����ҵ��˹̹����ѧ�������ݲ�ʿ��1974��ؼ����������������ڣ�������������ʱδͨ������Ҫԭ��������̫�١�����Clarkson��ѧ�ν̣������±䶯���ٴ�δ���������ڣ�1990��ص����ݸ�����������2015�����ಡȥ����Rosbash��2015����
�����������
���죬�����ӵ���Ҫ���Ⲣδȫ�������
�Ѿ����ֵIJ��������ӵĻ��������������ȫ���������磬�о�����ࡢʱ�������Per��������ʮ�����Per��������ε��ڻ���ת¼�����磬Pt��cek��Fuʵ���Ҵ�2001��2005���о�����hPer2��S662λ�����ữ����Ҫ�������CKI����������һλ��ĵ���ø�������˾��ȵ��ǣ����������ʵ������2007�귢����һ����������CKI�ĵ���ø���������ữhPer2�����ʵ�S662��Xuet al.,2007������ˣ��Դ�2001�귢�ֵĶ��˵�������������Ҫ��S662λ�㣬��Ȼ����֪���䵰��ø��17��������ͬʵ����Ŭ���Թ���ʶ�����ữS662λ��Ŀ��壬�Ա�����ټ��S662�Ƿ����ữ�������ɹ��ı�����Per�������ж�����Ա����ữλ�㣬����ȷʵ�����ڱ����ữ�����Ǹ���ʲô�������壿
�˽���������ӵ����ʣ��ܷ��ܺõ���ѧģ�ͣ��ڶ��ԵĻ���������������ʱ����
����ϸ���ڵĻ������û��������ǻ��С�
���ӵ�ϸ�����������ã�ֻ����ϸ�����������ӣ�����������ϸ�����루Tso et al., 2017������·��ι�����
����������ε��������ӣ������ڹ�Ӭ���Dz����࣬������˵ʮ���������Ӭ�ⷽ���һ���о����ı����塣������ʵ�������С�������ͨ���������¶�Э�������ӣ�Ҳ��һ�����ձ�Ϊ���������ܡ�
���������л����л�����Ĺ�ϵ���������Turek et al., 2005��Lamia et al., 2008; Marchevaet al., 2010��Robles, Humphrey and Mann��2017��
�������۵������ӣ���Ҫ�ǽ�24Сʱ����ҹ���ɡ����з�24Сʱ���������ɣ����ǵĻ�����ʲô��
�����о�֮�⣬�����ӵ��о�Ӧ�ÿ��Բ�������Ӧ�õijɹ������������о�����û�ó�����ʱ���ҩ��ִ������ڵ���ʱ���нϴ�������ֵ�ർ��Ӣ��������֮һ���ϰ�����Ҫ��ʱ����������ʱ��������ÿ�������ڼƣ�������˯�������൱�������������쳣���£����Ե���ʱ����ڹ���Ч�ʺ���������������Ҫ����ҩ�����ؽ�������������������֢��������Ⱦ����������ʱ����ӿ�����ԭ��֮һ��
����������������ص�����������Ҫ�ģ����������ⲻ���˯�ߡ����������ӿ��Խ���˯�ߵ���λ����ʱ˯���������ܽ���˯�߱�����˯�ߵĻ���Ŀǰ������١��̿�����˯�ߵ������൱�����Ե�ͼ����棬��������������ǻ��������⡣˯���о����ص�Զ��Orexin����������о�����Ϊ��ͻ�ƣ��������Ľ�չ���ޡ���2016����?��ʷ��MasashiYanagisawa����������߷��ֵ�Sik3�����Ƿ���˯�߷��ӻ����Ĺؼ�ͻ�ƣ�ֵ����Ŀ�Դ������꣩
����ע��������Դ���ڿΣ�����ʼ��2014��10��6�գ�������2015��2016��10�£���һ�������2017��10��1�գ�10��2�ղ����ġ�
ע1����Ӭ���Ĵ�ŵ������Ϊ1933��������Ħ�������ḻȾɫ���Ŵ�ѧ˵���ɣ���1947��Ħ������ѧ�����գ�����X���յ�����ͻ�䣩��1995��������·��˹���¹���ť˹��-�ֹ��ء�������ά����˹�������Ļ���2011�귨���Ļ��������������ߵĻ���
ע2��ϣ����������������������������Ч�����ߡ�����������������õ����²���һǧ���ٶ�Σ���ͬ���������Ϻõ�����һ�����ü��ٴΡ����������ֱ�һЩ���������ٺܶࡣ��ʵ��˵���������ƶ�����Ҫ���о��������������ö����������������ڡ��������ؼ���Ҫ�о����˶ࡣ
ע3����ȵĽ���Ϊcirannual����ҹ���ӽ�24Сʱ��Ϊcircadian������24Сʱ��Ϊultradian������24Сʱ��Ϊinfradian��
ע4��JohnWatson���о������飬��һ�����������ϰ��Ļ��ߵó������ܷ��ƹ㣬����������������С����������ʵ�����������С���IJ���Ӱ�졣
ע5������ʡ���˶�Konopka��Benzer��attached X�IJ��֡�Ħ����������LilianMorgan�����Ƕ����Ŀ�ѧ�ң�������ͣ����ʮ���꣬������Ů���ط�ʵ���ң�ֻ����Morganʵ���ң�����ʮ���귢�ֵ�attachedX��Ӭ�����ڹ�Ӭ�о������ô���Konopka��Benzer����attached X��ʡ��һ���Ľ��䣬�ӿ���ɸѡ�ٶȡ�
ע6����Ȼֻ�ж������Ӿ����������ֲ��Լ���Щϸ�������ий�ϵͳ���������ֲ��ֱ���һЩ���Ӹй⣬��ֻ��Cry�������Ƕ�ֲ�ﶼ���ڸй�ķ��ӡ��Ȿ��Ҳ���о�Cry�Ƿ������ҹ���ɵ�һ��ԭ����ֵ��ǣ�������ʵ�����������ҹ���ɵ�Cry�����ܸй⣬����Cry�����û���������
��л����л���������Ŷ�������謡���ǿ����������������ϣͬ���Ļ����� |

















